




Дьяченко И.П. Фауна и некоторые биологические особенности рыб в эпоху поздней бронзы
Одним из способов ретроспективной оценки ихтиофауны является изучение кухонных остатков в культурных слоях поселений человека минувших эпох. Конечно, такой метод не лишен
100
некоторых недостатков, связанных прежде всего с селективностью рыболовства, имеющей в своей основе те или иные причины. Но, с другой стороны, именно эта избирательность способна оказать содействие в разрешении некоторых крайне важных вопросов о направлениях развития хозяйственной деятельности наших далеких предков, характере и уровне их воздействия на состояние и динамику естественных экосистем. Кроме того, такие данные могут быть использованы для конкретизации наших представлений о темпах эволюции групп, органов и функций. Наконец, полученные результаты в ряде случаев могут послужить основанием для корректировки тактических подходов к проблеме охраны и рационального использования рыбных запасов во внутренних водоемах.
Материал и методика исследований
Исходным материалом для выполнения настоящей работы послужили коллекции чешуи нескольких видов пресноводных рыб, составленные работниками совместной экспедиции Башгосуниверситета и Башгоспединститута под руководством М. Ф. Обыденнова и В. С. Горбунова, проводившей археологические раскопки Юкалекулевского поселения, расположенного на берегу р. Ай в пределах Кигинского района БАССР. Вся обнаруженная чешуя была предоставлена в наше распоряжение.
Датировка поселения проведена М. Ф. Обыденновым по находкам материальной культуры, указывающим на то, что они относятся к концу II тыс. до н. э.
Примечательно, что на местах раскопок не обнаружено мощных скоплений чешуи и костей рыб, характерных для постоянных мест обработки рыбы у славянских племен, населявших Европейскую часть территории СССР, у которых рыболовство было хорошо развито в неолите[1], а также у племен, селившихся в X—XII вв. до н. э. на Оби[2]. В Юкалекулевском поселении чешуя рыб была найдена только в двух местах и в ограниченном количестве. В то же время последующий анализ показал, что наш материал, состоящий всего из 307 экземпляров целых чешуек и их поддающихся диагностированию фрагментов, крайне разнообразен. Эти остатки принадлежали особям 12 видов рыб семейства карповых, что говорит, скорее всего, о длительном использовании мест обработки рыбы. Кроме всего прочего, в этом факте заключена реальная основа для предположения, что у южноуральских племен во времена срубной и межовской культур бронзового века рыба не играла существенной роли в пищевом рационе. Для сравнения можно привести данные А. Н. Гундризера[3], обнаружившего в поселении Еловка на Оби более 900 г, т. е. несколько тысяч, чешуи, принадлежавших всего 4 видам рыб, а преимущественно язю и карасю золотому.
Чешуя рыб, которой мы располагали, имела различную степень сохранности и фоссилизации. Обыизвествление чаще всего затрагивало центральную часть чешуйного поля, затрудняя
101
определение возраста, поэтому в ряде случаев точность возрастной диагностики была на уровне не более +1—2 года.
В связи с отсутствием в находках костных остатков точные размеры тела рыб установить не представляется возможным. О них можно судить лишь приближенно, ориентируясь на величину чешуйных пластинок, так как известно, что во многих случаях имеет место прямая корреляция между линейными размерами тела рыб и размерами их чешуи[4][5]. Однако здесь возникает новая проблема, потому что требуется установить, к какой части тела относится ископаемая чешуя, так как ее структура, в том числе и размеры в разных частях тела, неодинакова, как и у современных рыб. Значит, если принять во внимание, что величина чешуйных пластинок на определенном участке тела строго соответствует определенной длине тела и соотношение между этими величинами у всех особей вида остается постоянным (а это не всегда так), то и тогда пришлось бы перебрать сотни эталонных препаратов от каждой особи, прежде чем удалось бы найти аналог ископаемой чешуйке. Но в принципе использование подобного метода реставрации линейных размеров тела, основанного на идентификации чешуи, в определенных случаях может стать рациональным.
При анализе ископаемого материала мы опирались на распространенное мнение о том, что в среднем морфобиологические признаки рыб четвертичного периода практически не отличаются от таковых у рыб современности[6]. В этой связи очень интересным является представление о том, что не только морфобиологические показатели рыб, но и фаунистический облик вплоть до уровня вида, сходный с современным, сформировался уже на границе плиоцена и плейстоцена, т. е. непосредственно между третичным и четвертичным периодами кайнозоя[7]. Более того, В. Д. Лебедев[8] считает, что этот процесс совершился еще раньше — где-то в глубине плиоцена или не менее 3—4 миллионов лет тому назад, если придерживаться геохронологической шкалы, приведенной А. В. Яблоковым и А. Г. Юсуфовым[9].
Немаловажное значение в свете стоящих задач имеет проблема ретроспективной оценки внутригодовых сроков вылова рыб, которую мы попытались решить путем анализа структуры чешуи, в частности характера расположения так называемых годовых колец и относительной ширины зоны краевого прироста. Здесь было принято во внимание классическое мнение, разделяемое многими ихтиологами, о том, что образование нового годового кольца на чешуе происходит в начале нового периода роста — поздней весной или в начале лета у неполовозрелых особей или даже во второй половине лета, что часто наблюдается у половозрелых рыб[10].
Видовая диагностика ископаемых рыб по чешуе была проведена с помощью серии эталонных препаратов чешуи различных видов современных рыб под контролем фотографий чешуи, приведенных Г. Г. Галкиным[11] в Атласе чешуи пресноводных
102
рыб СССР. В качестве основных диагностических признаков принимались пропорции чешуи, топография склеритов, количество, характер расположения и рисунок радиальных каналов, расположение образовательного центра, а также конфигурация орального, каудального и латерального краев чешуйной пластинки. Некоторыми из этих признаков мы успешно пользовались и раньше[12].
Таким образом была установлена видовая принадлежность 146 чешуи из 307 имевшихся в нашем распоряжении (47,6%) (см. табл. 1).
Вопрос определения возраста ископаемых рыб по чешуе или ее фрагментам оказался более сложным в силу ряда объективных причин. Главными из них были следующие:
1. Отсутствие у некоторых чешуек внутренних годовых колец из-за резорбции центральной поверхностной части — весьма обычное явление и у современных рыб, хотя его причина по сей день остается загадочной.
2. Фоссилизация центрального ядра некоторых чешуи, сопровождающаяся разрушением части склеритов.
3. Некоторые фрагменты чешуи не охватывали целого радиуса, в результате чего терялись внутренние или наружные годовые зоны.
Кроме того, встречались отдельные чешуйки с гомогенной склеритной структурой, что известно у отдельных популяций современных рыб.
Просветление чешуи проводилось их выдерживанием в 3%-ом растворе соляной кислоты или в глицерине в течение длительного срока. Поверхность чешуи очищалась мягкой щеточкой. Всего возраст был определен по 139 чешуям (табл. 1).
Таблица 1
Объем чешуйного материала из Юкалекулевского поселения
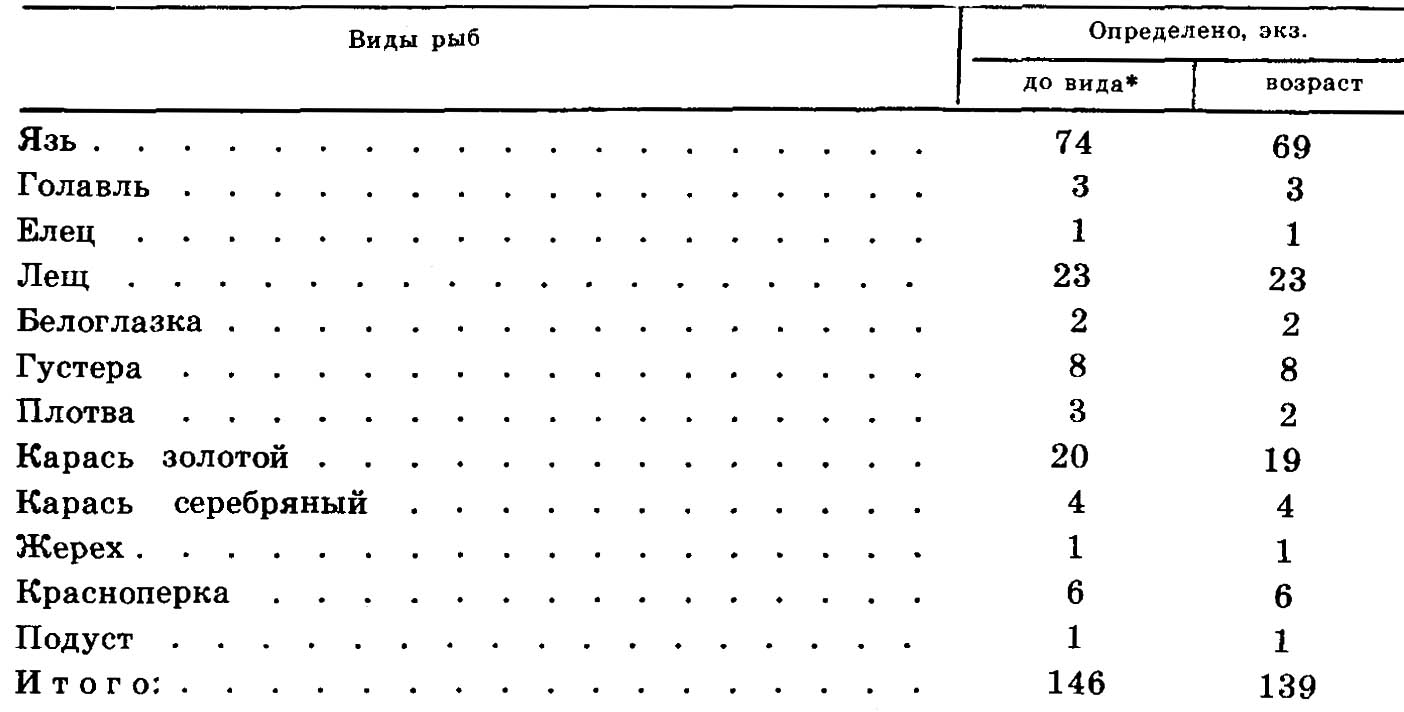
*Кроме того, до уровня семейства определена 161 чешуя. Вся она принадлежит видам сем. карповых.
103
Видовой и размерно-возрастной состав уловов рыб у человека бронзового века
Наиболее существенной и в известной мере загадочной чертой качественного состава уловов обитателей Юкалекулевского поселения, расположенного на одном из крупнейших притоков р. Уфы — реке Ай, является полное отсутствие представителей других таксонов, кроме рыб, принадлежащих к сем. карповых (табл. 1). Этот факт пока не может быть удовлетворительно объяснен, ибо никак нельзя допустить, что в этой реке, относящейся по существующей классификации к разряду средних рек[13], во времена бронзового века истории человечества не было другой рыбы, кроме видов сем. карповых, чьи остатки были обнаружены археологами. Ведь в верховьях реки Камы, т. е. в климатически и гидрологически сходных условиях, обитали кроме видов сем. карповых еще и рыбы из семейств осетровых, лососевых, щуковых, сомовых, тресковых и окуневых[14][15]. Правда, ископаемые свидетельства в виде костей и чешуи, найденные среди кухонных остатков, относятся к более позднему времени — X—XII вв. н. э., однако, существенного изменения климата, способного повлечь за собой кардинальную перестройку экосистем рек, за последние несколько тысячелетий не произошло (Никольский, 1935— цит. по Лебедеву)[16], а уровень антропогенного воздействия в условиях разреженного населения в обе рассматриваемые эпохи должен быть вполне соизмеримым. Таким образом, оснований для предположения о резкой смене ихтиокомплексов за этот период нет, так как, в соответствии с данными ряда авторов, их современный вид в бассейнах Белого, Балтийского, а тем более Каспийского, Азовского и Черного морей сложился уже, самое позднее, «между концом II и началом I тысячелетия до н. э.»[17] Вот и в реках Ай, Юрюзань и Уфе, протекающих в одной климато-географической зоне, сейчас, как показывают наши наблюдения, встречаются представители всех перечисленных для верховьев р. Камы семейств класса костных рыб.
Таким образом, факт селективного отлова карповых рыб общиной Юкалекулевского поселения можно назвать бесспорным. Однако решить вопрос чем это было обусловлено, непросто. Не исключено, что местное население, являющееся по существующему мнению генетически связанным с ираноязычными племенами[18], в соответствии со своими культами, верованиями оставляло в покое хищников (щука, сом, окунь), а тем более налима, нередко питающегося мертвечиной. К тому же совершенно очевидно, что, как и у срубных племен (середина II тыс. до н. э.)[19], рыболовство играло незначительную роль в хозяйственной деятельности. Об этом свидетельствует хотя бы то, что члены Юкалекулевского поселения добывали рыбу преимущественно в наиболее доступное для промысла время — весной в период нереста и следующие за ним несколько летних месяцев (табл. 2).
104
Таблица 2
Внутригодовое распределение уловов рыбы
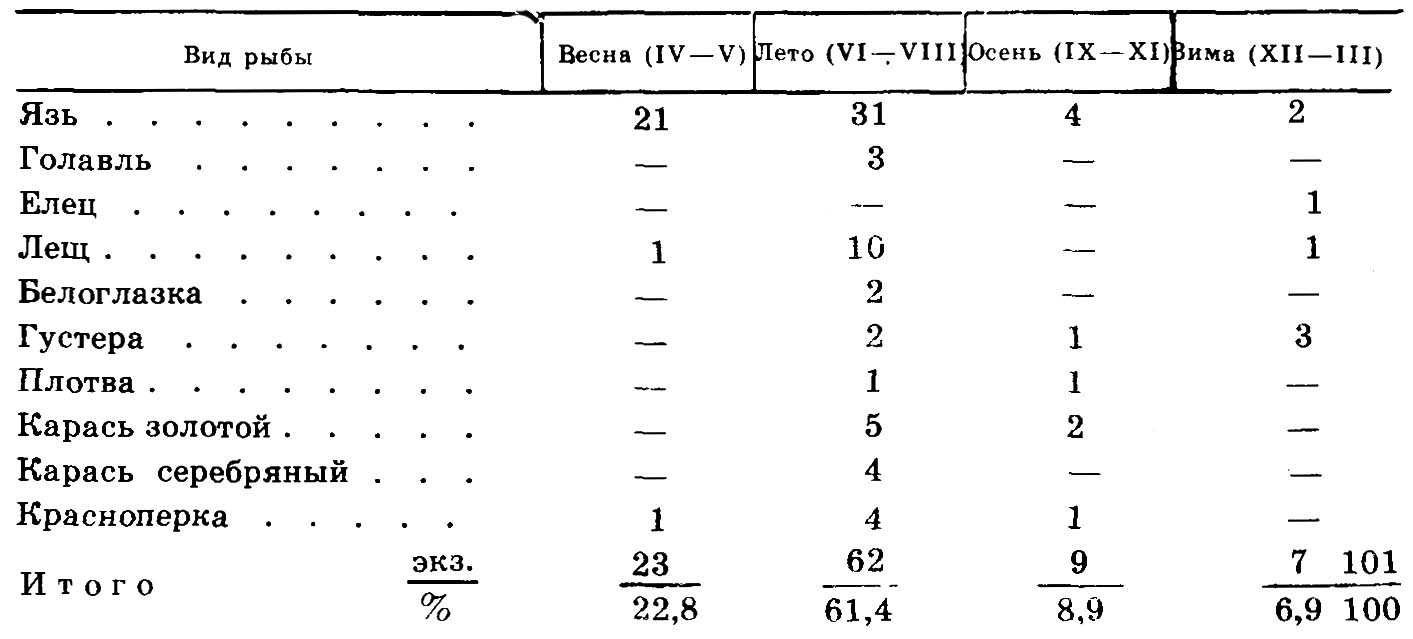
Из поддающегося хронологизации чешуйного материала (101 экз.) видно, что на весенне-летний период приходится 84,2% всего улова рыб. Эта диспропорция внутригодового распределения уловов лишь подтверждает мнение, что рыболовство было побочным занятием местного населения. К тому же для отлова рыб оно не пользовалось, видимо, специальными орудиями (отцеживающие, крючные), так как при раскопках не было обнаружено сетных или неводных грузил, острог, крючьев, столь характерных среди предметов, добываемых археологами в поселениях славянских племен той же эпохи[20].
Добыча рыбы велась какими-то примитивными способами в период их массовых скоплений, когда не требуется прибегать к особым ухищрениям. Успеху в промысле способствовало также обилие рыбы, а в пищу употреблялись главным образом крупные экземпляры, относящиеся к старшим возрастным группам (табл. 3).
Материалы табл. 3 показывают, что по наиболее широко представленному в уловах язю промысел базировался главным образом на 7—11-годовалых особях, по стоящему на 2 месте лещу — на 10—12-годовалых, по карасю золотому — на 6—12-годовалых, а остатки всех 3 экз. плотвы, а также жереха и поду ста принадлежали особям старше 10 лет. В целом по всему контингенту вылова 89,3% рыб имели возраст от 7 до 15 лет и лишь 10,7% были в возрасте от 4+ до 6+ . Чешуя более молодых рыб не обнаружена.
Таким образом, наши материалы отражают тенденцию базирования промысла по основным видам рыб (лещ, язь, голавль, белоглазка, плотва, карась) на старшевозрастные группы. Они фактически совпадают с данными, приведенными в монографии В. Д. Лебедева (1960), в статьях А. И. Букирева, Э. А. Усольцева (1958), А. Н. Гундризера (1966) и многих других.
105
Таблица 3
Возрастной состав рыб в промысловых уловах у человека бронзового века

104
Характер роста рыб
Степень антропогенного воздействия на природные популяции животных во многом определяет их биологическое состояние, в частности, динамику численности, темп роста всех или отдельных возрастных групп. Это и понятно, ибо как численность, так и темп роста зависят, прежде всего, от кормовой обеспеченности. Поскольку темп роста и плотность населения рыб при стационарности корцовых ресурсов водоема находятся в обратной связи, то интенсификация промысла или иные формы антропогенного пресса, вызывающие снижение численности, способны вызвать повышение темпов роста, в то время как запуск рыболовства сопровождается обратными процессами. Поэтому неудивительно, что интенсивность роста ископаемых рыб — в Сибири или на Урале, в Приднепровье или в Поволжье — в условиях разреженного населения и несовершенства орудий лова, применяемых человеком прежних эпох, была, как правило, значительно ниже, чем у современных рыб[21].
Не составляет исключения и наш материал, хотя при отсутствии костных остатков очень сложно установить размеры рыб, чтобы затем, используя известные в ихтиологии методы, восстановить темп роста по какой-либо регистрирующей структуре, к примеру, по чешуе.
Конечно, достаточно объективные данные о темпах роста сравниваемых рыб может дать и непосредственное сопоставление ширины годичных колец. Однако эти годичные зоны на чешуе из различных участков тела, а иногда даже из идентичного участка разных особей исследуемой популяции, могут различаться[22]. Вместе с тем, вероятность крупных ошибок при сравнительной оценке темпов роста по аналогичным чешуям будет значительно ниже, поэтому очень важно правильно определить локализацию ископаемой чешуи на теле. Нивелирование возможных ошибок достигается также использованием средних значений, ибо «по средним размерам рыбы и соответствующим им средним размерам чешуи ... обнаруживается хорошо выраженная связь»[23]. Все это и понятно, так как в данном случае устраняются помехи индивидуальной изменчивости роста, характера корреляции в интересующей нас системе.
Идентификация ископаемых чешуи проведена на основании того, что их форма, относительные размеры пластинки, те или иные топографические детали в целом вполне определенно, закономерно изменяются в зависимости от места расположения на теле[24]. Учитывая это, нам удалось идентифицировать чешую от 5 особей ископаемого леща и 4 особей язя. Для сравнения показателей роста мы использовали аналогичные чешуи от 6 экз. леща озера Шамсутдин, расположенного в пойме р. Белой (среднее течение), ежегодно заливаемого в половодье, а также 7 экз. язя из Павловского водохранилища на р. Уфе. Возраст сравниваемого леща был в пределах 6—9 лет (современный) и
107

Рис. 1. Темп роста ископаемых и современных язя и леща по данным измерений ширины годовых зон на чешуе (в делениях окуляр-микрометра при увеличении 8×1)
10—11 лет (ископаемый), а язя, соответственно 4—5 и 7—8 лет. Измерение радиусов чешуи проводились в направлениях, по которым наиболее отчетливо прослеживались годичные кольца (рис. 1).
Результаты анализа полученного материала оказались неодинаковыми по двум сравниваемым видам. Выяснилось что темп роста ископаемого леща, как и ожидалось, в целом значительно уступал интенсивности роста нынешнего леща. Однако вызывает интерес то обстоятельство, что заметное отставание в темпе роста начинается только с третьего года жизни и в
108
последующем более высокая интенсивность роста современного леща выглядит бесспорной (рис. 1).
Среди ряда возможных причин такого явления наиболее реальной можно считать ту, что обеспеченность пищей, а, значит, и численность молоди леща до трехлетнего возраста была одного порядка, что и в настоящее время. Этим определяется сходство темпов роста молоди. В последующем показатели роста расходятся, что является отражением изменения характера воздействия среды: с возрастанием интенсивности промысла улучшается кормовая обеспеченность эксплуатируемой части стада леща.
Иначе выглядит результат сравнительного анализа роста язя. Оказалось, что средние значения радиусов на чешуях современного и ископаемого язя за соответствующие годы жизни различаются совсем мало, это на графике выражается почти идеальным совпадением линий регрессии. По аналогии с логикой вышеизложенных рассуждений можно прийти к выводу, что совпадение темпов нарастания чешуи (а значит и тела) у сравниваемых выборок язя должно быть прежде всего следствием идентичности комплекса воздействующих факторов, в том числе промысла. Другие компоненты этого комплекса, например, климат, гидрологический режим, вряд ли существенно изменились за 3 тыс. лет, разделяющих наши эпохи. Такого заключения нельзя сделать относительно уровня воздействия антропогенного фактора, главным образом интенсивности промысла, если исходить из показателей плотности народонаселения, сопоставления эффективности методов и орудий рыболовства, которые бесспорно уступали современному уровню. Тогда в чем заключается причина обнаруженного эффекта?
Вполне реальным является предположение о значительной интенсивности вылова язя людьми бронзового века на Южном Урале, в результате чего численность этого вида рыб была сопоставима с современной. Косвенным доказательством такого положения можно считать то, что чешуя язя в кухонных остатках обитателей Юкалекулевского поселения наиболее многочисленна. Она составляет более половины обнаруженных здесь и определенных до вида чешуек (табл. 1). Этот факт может служить также дополнительным аргументом в пользу того, что наши предки избирательно относились к промысловым объектам.
Таким образом, материалы проведенного анализа свидетельствуют о том, что темп роста рыб в одной климато-географической зоне и в условиях одинаковой промысловой эксплуатации их популяций имеет сходные абсолютные величины и такую же биологическую специфику, во всяком случае на протяжении не менее 3 тыс. лет. Это указывает на то, что характер роста в реальных условиях является одним из важных видоспецифических свойств рыб, а также подтверждает правомерность сравнительного анализа рыб неолита и современности.
109
Кроме того, определяется очень важное следствие практического характера. Становится очевидным, что популяции леща в бассейне р. Белой сейчас подвергаются промысловому воздействию, начиная уже с трехлетнего возраста при длине тела 18,0—23,0 см, т. е. задолго до наступления воспроизводительной способности. Если учесть, что к размножению лещ у нас приступает в возрасте 5—6 лет, а то и позже, достигнув размеров, как правило, не менее 28,0—32,0 см, станет ясно, что систематическим выловом неполовозрелого леща наносится ощутимый ущерб воспроизводству основного промыслового объекта в нашей республике.
ЛИТЕРАТУРА
[1] Лебедев В. Д. Пресноводная четвертичная ихтиофауна европейской части СССР. М.: Изд. МГУ, 1960.—404 с.
[2] Гундризер А. Н. Рыбы из поселения Еловка на Оби.— В кн.: Ученые записки, № 60. Томск: Изд. Томск, ун-та, 1966, с. 119—123.
[3] Там же.
[4] Врюзгин В. Л. Методы изучения роста рыб по чешуе, костям и отолитам. Киев: Наукова думка, 1969.—187 с.
[5] Яковлева А. С. Использование особенностей структуры чешуи рыб при экологическом анализе.— В кн.: Морфоэкологические особенности рыб бассейна реки Северной Сосьвы. Свердловск: Изд. УНЦ АН СССР, 1979, с. 86—93.
[6] Лебедев В. Д. Указ. соч., с. 373.
[7] Thinemann А. Die Binnengewässer. Verbreitungsgeschichte der Tierwelt Europas. — Stuttgart: Bd. 18, 1950. — 809 s.
[8] Лебедев В. Д. Указ. соч.
[9] Яблоков А. В., Юсуфов А. Г. Эволюционное учение. М.: Высшая школа, 1981.—343 с.
[10] Никольский Г. В. Экология рыб. М.: Высшая школа, 1974.—367 с.
[11] Галкин Г. Г. Атлас чешуи пресноводных костистых рыб.—Изв. ВНИИ озерного и речного рыбн. хоз-ва, т. 46, 1958.
[12] Дьяченко И. П. Использование топографических деталей чешуи рыб при определении их возраста.— В кн.: Видовой состав и экология животных Башкирии. Уфа, 1976, с. 135—142. (Рук. деп. ВИНИТИ 1 июня 1976 г., № 1974 — 76 Деп.).
[13] Чеботарев .4. И. Гидрологический словарь. Л.: Гидрометеоиздат, 1978.—308 с.
[14] Букирев А. И., Усольцев Э. А. К истории ихтиофауны бассейна реки Камы.— Зоологический журн., т. 37, вып. 6, 1958, с. 884—898.
[15] Лебедев В. Д. Указ. соч.
[16] Там же.
[17] Там же.
[18] Генинг В. Ф. История населения удмуртского Прикамья в пьяноборскую эпоху.—ВАУ, вып. 10, 1970, с. 18.
[19] Там же.
[20] Лебедев В. Д. К вопросу об изменении ихтиофауны реки Десны в период от последней межледниковой до современной эпохи.—Зоологический журн., т. 23, вып. 5, 1944, с. 240—249.
[21] См.: Лебедев В. Д. К вопросу об изменении ихтиофауны реки Десны...; он же. Пресноводная четвертичная ихтиофауна европейской части СССР; Букирев А. И., Усольцев Э. А. Указ. соч.;Г ундризер А. Н. Указ. соч.; и др.
[22] Брюзгин В. Л. Указ. соч.
[23] Там же, с. 37.
[24] Дьяченко И. П. О функциональных основах роста и некоторых структурных особенностях чешуи рыб.— В кн.: Материалы по фауне водоемов Башкирии, вып. 76, № 9. Уфа: Изд. Башк. ун-та, 1975, с. 51—62.
110
ПУБЛИКАЦИЯ: Дьяченко И.П. Фауна и некоторые биологические особенности рыб в эпоху поздней бронзы // Бронзовый век Южного Приуралья: Межвузовский сборник научных трудов. Уфа, 1985. С. 100-110.