




Цепкин Е.А. Остатки рыб из пещеры Кударо I
Изучение четвертичной фауны рыб по костным остаткам из раскопок археологических памятников представляет интерес для археологов, ихтиологов, палеоклиматологов и специалистов некоторых других смежных дисциплин. Трудами отечественных исследователей за последние, десятилетия накоплен большой материал, характеризующий ту фауну рыб эпох неолита, бронзы, железа, которую промышлял человек. Для ряда водоемов выяснен видовой состав фауны, восстановлены некоторые биологические показатели субфоссильных рыб. Однако сведения о фауне рыб в палеолите чрезвычайно скудны. В палеолитических слоях остатки рыб встречаются редко и обычно имеют настолько плохую сохранность, что по ним даже определить вид рыбы удается далеко не всегда.
Редкие находки костей рыб в палеолитических стоянках и плохая сохранность их объясняют то, что ихтиофауна палеолита до сих пор очень слабо изучена. Естественно, каждая новая находка остатков рыб в культурном слое стоянок палеолитического человека представляет для исследователей исключительно большой интерес.
Материалом для настоящей статьи послужила обширная коллекция костей рыб, собранная В.П. Любиным в 1956—1959 гг. при раскопках пещеры Кударо I. Коллекция содержала 36537 костей, представленных многочисленными обломками плавниковых лучей, radialia и других частей осевого скелета, также позвонками, hypuralia и обломками костей головы (рис. 1, 2). Состав исследованных костных остатков и распределение их в плане и по слоям приведены в табл. 1—8.
Наибольшее количество остатков рыб отмечено в мустьерских культурных слоях восточной галереи, причем главное средоточие находилось в южной части этой галереи и повсеместно — в мустьерском горизонте 3 в. В ашельском же культурном слое больше всего костей встречено в центральной камере (раскоп 1959 г.).
При обработке ископаемого остеологического материала для определения вида рыбы используются, как известно, не все части скелета, а лишь те, которые обладают достаточно достоверными диагностическими признаками. В данной коллекции таковыми были позвонки, hypuralia, фрагменты челюстных и других костей головы. Все они принадлежали одному виду — черноморскому лососю Salmo trutta labrax Pallas. Отсутствие среди указанных определимых костных остатков костей других видов рыб дает полное основание считать, что мелкие фрагменты плавниковых лучей, radialia и других частей осевого скелета также принадлежат лососю.
По наиболее хорошо сохранившимся туловищным позвонкам и hypuralia удалось восстановить размеры рыб; они колеблются от 50 до 130 см (см. рис. 2; рис. 3)[1]. Современный проходной черноморский лосось, поднимавшийся на нерест в реки Кавказа, в первой половине нашего сто-
90

Рис. 1. Схематическое изображение остатков лосося. r — radialia; l — плавниковые лучи; v — позвонки; h — hypuralia
летия имел длину 35—120 см (в среднем 89 см). В кавказских реках встречается также мелкая пресноводная форма — форель, размеры ее 10—24 см, в среднем 16 см [Барач, 1962]. В коллекции из пещеры Кударо I среди костей крупных лососей попадались фрагменты костных остатков, принадлежавших особям длиной около 20—40 см. Видимо, это костные остатки пресноводной жилой формы лосося — форели.
При сравнении размеров лосося из разных слоев пещеры Кударо I отмечено уменьшение средней и максимальной длины субфоссильных рыб от нижнего ашельского слоя 5 к верхнему мустьерскому горизонту 3а (табл. 9).
Изменение биологических показателей рыб происходит, как известно, под воздействием антропогенных (промысел) и физико-географических факторов. В результате интенсивного вылова в ряде водоемов уменьшились средние размеры промысловых рыб (при этом максимальная длина их почти не изменилась).
Вряд ли можно допустить, что рыболовство в среднем палеолите было столь интенсивным, что оказало заметное влияние на размерный состав популяции лосося р. Риони. Палеоихтиологические исследования показывают, что заметное воздействие промысла на фауну рыб континентальных водоемов стало проявляться лишь в последние два-три столетия [Лебедев, 1960; Цепкин, 1966].
Уменьшение средних и максимальных размеров черноморского лосося от ашеля до мустье, несомненно, следует отнести за счет влияния физико-географических условий (изменение климата, гидрологического режима кавказских рек и других факторов, не связанных с хозяйственной деятельностью человека).
На некоторых наиболее хорошо сохранившихся позвонках лосося из пещеры Кударо I заметны годовые кольца: первые два-три — сравнительно узкие, последующие — широкие[2]. Расположение годовых колец на позвон-
91
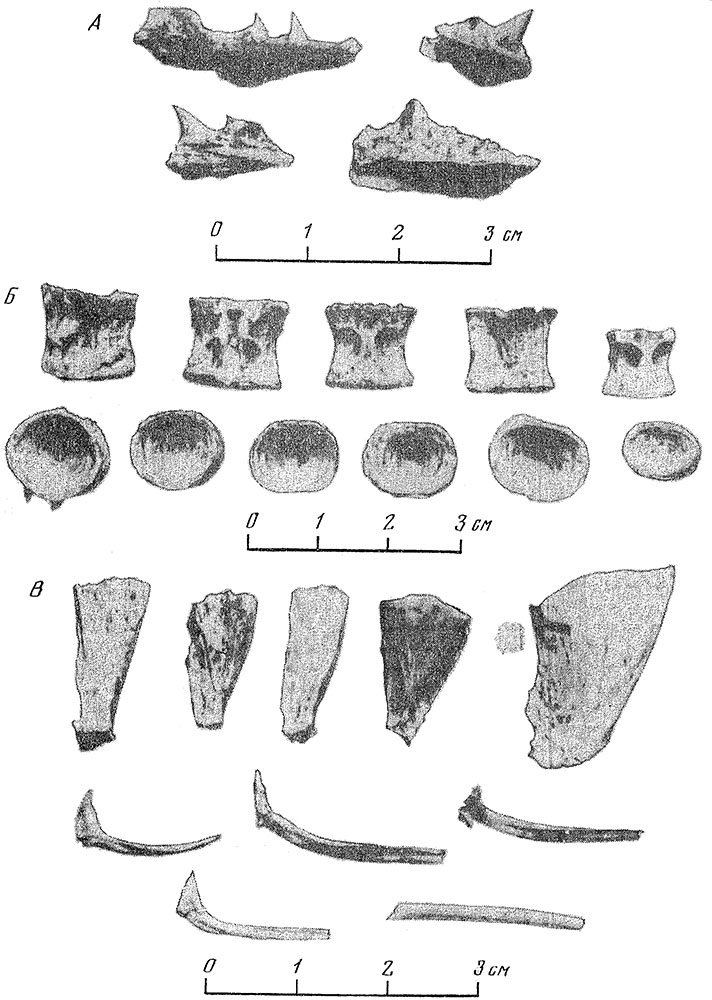
Рис. 2. Костные остатки лосося из пещеры Кударо I. А — фрагменты челюстных костей; Б — позвонки; В — hypuralia и обломки плавниковых лучей
92
Таблица 1
Остатки рыб (36537 экз.) в мустьерских и ашельских слоях на участках, раскопанных в 1956—1959 гг.
|
Участки |
Мустьерские слои 3, 4 |
Ашельский слой 5 |
Всего |
|
|
Южная галерея, 1956 |
— |
12 |
12 |
|
|
Восточная галерея |
|
|
||
|
северная часть, 1957 г. |
2968 |
32341 |
||
|
южная часть, 1958 г. |
28226 |
1147 |
|
|
|
Центральная камера, |
|
|
||
|
восточная часть, 1959 г. |
2867 |
1317 |
4184 |
|
ках наряду с крупными размерами самих рыб позволяют отнести этих лососей к крупной проходной форме Salmo trutta labrax Pallas.
Проходной черноморский лосось и форель были обнаружены М.И. Тихим в остатках рыб из крымских пещер Сюрень I (ориньяк) и Сюрень II (поздний азиль) [Тихий, 1929]. Костные остатки лосося определены Г.В. Никольским в коллекции из Новгород-Северской палеолитической стоянки, В.Д. Лебедевым — из позднепалеолитической стоянки Мурзак-Коба в Крыму [Никольский, 1952; Лебедев, 1952].
Эти данные в совокупности с материалом из раскопок пещеры Кударо I свидетельствуют о широком распространении лосося рода Salmo в бассейне Черного моря в плейстоцене — раннем голоцене.
В настоящее время проходной черноморский лосось встречается главным образом у побережья Абхазии; в устье Риони и южнее он редок. Жилая форма его (форель) населяет многие горные реки и озера Кавказа [Барач, 1962].
Особо следует рассмотреть вопрос о том, что представляют собой многочисленные кости лосося, как они оказались в культурном слое пещеры Кударо I.
Н.К. Верещагин, исследовавший фауну из раскопок пещеры Кударо I, в монографии, посвященной млекопитающим Кавказа, пишет: «Обилие костей рыб в нижнепалеолитических слоях обоих входов пещеры Кударо I наводит на мысль, что отложения образованы волнами какого-то водоема, возможно пра-Джоджори, приносившими трупы и истиравшими кости в гальке и песке» [Верещагин, 1959].
С этим выводом Н.К. Верещагина трудно согласиться. Результаты исследования остеологической коллекции убеждают нас в том, что многочисленные кости рыб, найденные при раскоп-
Таблица 2.
Состав костных остатков из южной галереи. Раскоп 1956 г., ашельские слои
|
Кости |
Количество экз. |
|
Позвонки |
1 |
|
Плавниковые лучи |
11 |
|
Всего |
12 |
Таблица 3
Состав костных остатков из восточной галереи (северная часть). Раскоп 1957 г., мустьерские и ашельские слои
|
Кости |
Количество экз. |
|
Челюстные кости |
6 |
|
Кости головы |
89 |
|
Позвонки |
421 |
|
Hypuralia |
62 |
|
Плавниковые лучи, radialia и другие фрагменты осевого скелета |
2390 |
|
Всего |
2968 |
Таблица 4
Состав и распределение костных остатков (28226 экз.) из восточной галереи (южная часть). Раскоп 1958 г. Мустьерские слои.
|
Кости |
Количество экз. |
|||
|
Горизонт 3а |
Горизонт 3б |
Горизонт 3в |
Слой 4 |
|
|
Челюстные кости |
1 |
4 |
45 |
16 |
|
Кости головы |
32 |
27 |
326 |
38 |
|
Позвонки |
75 |
346 |
4402 |
543 |
|
Hypuralia |
7 |
26 |
205 |
40 |
|
Плавниковые лучи, radialia и другие фрагменты осевого скелета |
1253 |
2574 |
16499 |
1767 |
|
Всего |
1368 |
2977 |
21477 |
2404 |
Таблица 5
Состав и распределение костных остатков (2867 экз.) из центральной камеры (восточная часть). Раскоп 1959 г., мустьерские слои
|
Кости |
Количество экз. |
|||
|
Горизонт За |
Горизонт 36 |
Горизонт 3в |
Слой 4 |
|
|
Челюстные кости |
— |
2 |
— |
— |
|
Кости головы |
2 |
15 |
27 |
3 |
|
Позвонки |
25 |
79 |
181 |
51 |
|
Hypuralia |
10 |
17 |
23 |
3 |
|
Плавниковые лучи, radialia;n другие фрагменты осевого скелета |
483 |
602 |
1104 |
240 |
|
Всего |
520 |
715 |
1335 |
297 |
94
Таблица 6
Состав и распределение костных остатков (1147 экз.) из восточной галереи (южная часть). Раскоп 1958 г. Ашельский слой 5
|
Кости |
Условные раскопочные горизонты |
||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
|
|
Челюстные кости |
1 |
— |
— |
— |
— |
— |
— |
— |
— |
|
Кости головы |
— |
3 |
5 |
2 |
3 |
— |
3 |
— |
— |
|
Позвонки |
7 |
5 |
8 |
13 |
3 |
13 |
1 |
3 |
15 |
|
Плавниковые лучи, radialia и другие фрагменты осевого скелета |
27 |
74 |
144 |
170 |
114 |
146 |
69 |
179 |
139 |
|
Всего |
35 |
82 |
157 |
185 |
120 |
159 |
73 |
182 |
154 |
Таблица 7
Состав и распределение костных остатков (1317 экз.) из центральной камеры (восточная часть). Раскоп 1959 г. Ашельский слой 5
|
Кости |
Условные раскопочные горизонты |
|||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
|
|
Кости головы |
— |
— |
1 |
2 |
— |
4 |
3 |
— |
|
Позвонки |
5 |
5 |
6 |
5 |
9 |
14 |
11 |
5 |
|
Hypuralia |
— |
— |
— |
— |
— |
1 |
— |
— |
|
Плавниковые лучи, radialia и другие фрагменты осевого скелета |
50 |
168 |
185 |
177 |
80 |
323 |
242 |
21 |
|
Всего |
55 |
173 |
192 |
184 |
89 |
342 |
256 |
26 |
Таблица 8
Состав и распределение костных остатков (2464 экз.) в ашельском слое 5. Раскопы 1958-1959 гг.
|
Кости |
Условные горизонты |
||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
|
|
Челюстные кости |
1 |
— |
— |
— |
— |
— |
— |
— |
— |
|
Кости головы |
— |
3 |
6 |
4 |
3 |
4 |
6 |
— |
— |
|
Позвонки |
12 |
10 |
14 |
18 |
12 |
27 |
12 |
8 |
15 |
|
Hypuralia |
— |
— |
— |
— |
— |
1 |
— |
— |
— |
|
Плавниковые лучи, radialia и другие фрагменты осевого скелета |
77 |
242 |
329 |
347 |
194 |
469 |
311 |
200 |
139 |
|
Всего |
90 |
255 |
349 |
369 |
209 |
501 |
329 |
208 |
154 |
95

Рис. 3. Размеры лосося из пещеры Кударо I; длина, см. 1 — ашель: М=88,5+3,9 см; σ=18,7; n=23 экз; 2—5 — мустье: 2 —слой 3а, М=80 см; n=9 экз.; 3 — слой 3б, М=84,2±1,39 см; σ=9,7; n=49 экз.; 4 — слой 3в, М=87,7±0,75 см; σ=13,2; n=305 экз.; 5 — слой 4, М=90,1±1,5 см; σ=12,4 см; n=71 экз.
ках в палеолитических слоях пещеры Кударо I, — остатки лососей, которых на протяжении тысячелетий добывал древнейший обитатель пещеры — человек.
Не случайно (об этом свидетельствуют археологические материалы) именно лосось стал одним из первых объектов древнепалеолитического промысла. Ежегодные массовые нерестовые миграции из моря в верховья рек этих крупных (длиной до 130 см, весом до 24 кг), обладающих превосходными вкусовыми качествами рыб, несомненно, с самых древнейших времен привлекали внимание палеолитического человека.
Черноморский лосось мечет икру в холодных горных реках и речках Кавказа, на каменисто-галечниковых грунтах, на глубине 50—70 см [Шарашидзе, 1968 ].
Чтобы достичь мест нереста, он идет из моря вверх по рекам, скорость течения которых местами превышает 3 м/сек. Во время этой миграции лосось преодолевает участки реки с перепадом воды (пороги) высотою до 1 м. Современный лосось проходит, например, по р. Кодор и притокам расстояние более 100 км и поднимается, таким образом, на высоту около 1000 м над ур. моря. Причем в более полноводные реки идут более крупные по размерам и весу лососи.
Характерная особенность представителей рода благородных лососей (Salmo), в том числе черноморского, состоит в том, что, проделав длительную, требующую огромных энергетических затрат, миграцию и отнерестившись в верхнем течении реки, они не погибают, а спускаются по реке на откорм в море и через год-два снова идут на нерест и т.д.
96
Приведенные основные экологические особенности миграционно-нерестового цикла черноморского лосося вряд ли могут вызвать сомнение в том, что многочисленные кости крупных ходовых лососей, найденные в нижнепалеолитических слоях пещеры Кударо I, являются кухонными остатками человека, обитавшего в этой пещере.
Промысел крупных идущих на нерест лососей не требовал ни лодок, ни сетей, ни крючков. Обстоятельствами, облегчающими добычу этих рыб, были небольшая глубина в местах скопления лососей на нерестилищах, усталость рыб, проделавших большой и трудный путь в верховье горной реки.
Древнейшие обитатели пещеры Кударо I всего вероятнее кололи лососей на нерестилищах деревянными копьями и дротиками или просто били камнями. Могли применяться и примитивные ловушки, сплетенные из прутьев и подвешенные у водопада, через который прыгали, скатывались вниз и снова прыгали идущие на нерест лососи. Этот весьма примитивный способ ловли лососей применялся в прошлом коренными жителями Карелии и Кольского полуострова. Таким же способом добывали лососей и северо-американские индейцы [Clark, 1948].
Состав костей, найденных при раскопках во всех палеолитических слоях, типичен для кухонных остатков рыб из культурного слоя палео- и неолитических стоянок (большое количество позвонков, фрагментов костей осевого скелета и малое количество костей головы).
Можно предположить, что какую-то часть рыбы потребляли медведи. Однако остатки съеденных ими лососей не могли бы образовать таких скоплений и именно в самой пещере, куда медведь должен был переносить каждую пойманную им рыбу. На Дальнем Востоке, например, медведи потребляют лососей, погибших после нереста — «снёнку», реже ловят их, когда они плотными косяками идут на мелководье рек. При обилии корма медведи съедают у рыб икру и внутренности или только мягкую часть спины или отгрызают голову рыбы, но никогда не переносят рыбу от места поимки, а съедают ее тут же на берегу реки.
Относительно малое количество фрагментов костей головы среди прочих костных остатков лосося не может служить доказательством того, что это остатки пищи медведя. Вероятность сохранения костей головы лосося по сравнению с другими костями ничтожно мала. Следует учитывать и то, что головы лососей несомненно употреблял в пищу палеолитический человек, поскольку голова этих рыб состоит главным образом из хряща и мягкой ткани и имеет нежные тонкие кости.
Таблица 9
Размеры лосося из разных слоев пещерыКударо I
|
Слой и горизонт |
Длина, см |
||
|
средняя |
максимальная |
||
|
3а |
80,0 |
100,0 |
|
|
3б |
84,2 |
105,0 |
|
|
3в |
87,7 |
115,0 |
|
|
4 |
90,1 |
115,0 |
|
|
5 |
118,0 |
130,0 |
|
97
[1] Приведена принятая для измерения лососевых рыб длина, определяемая по расстоянию от вершины рыла до конца средних лучей хвостового плавника.
[2] Рост рыб происходит периодически (в одни сезоны года рыба растет быстрее, чем в другие), что отражается на отдельных костях, в частности — позвонках. Периоды наибольшего роста заметны на позвонках в виде широких колец, периоды замедленного роста — в виде узких.
ПУБЛИКАЦИЯ: Цепкин Е.А. Остатки рыб из пещеры Кударо I // Кударские пещерные палеолитические стоянки в Юго-Осетии. М., 1980. С. 90-97.